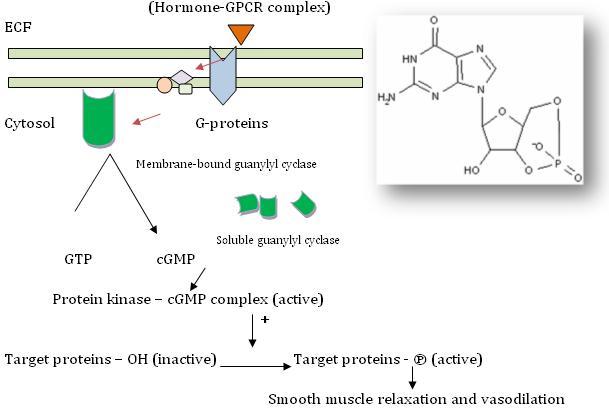
cGMP : A SECONDARY MESSENGER FOR ONE HORMONE AND A FEW COMPOUNDS
Cyclic guanosine 5’ – 3’ – monophosphate (cGMP) is made from guanosine – 5’ – triphosphate (simply, guanosine triphosphate) by the enzyme guanylyl cyclase, otherwise known as guanylate cyclase, which exists in cells in both membrane – bound and soluble (cytoplasmic) forms. Each of these isozymes differs in their kinetic, physiochemical and antigenic properties.
FUNCTIONS OF cGMP: -
Ø Atrial tissues in heart produce a group of peptides called atriopeptins (e.g. Atrial natriuretic factor [ANF]). These peptides that function as hormones, bind to membrane – bound guanylate cyclase and exert their action. There is a 50 fold increase in cGMP levels which is thought to mediate the effects such as natriuresis, diuresis, vasodilation and inhibition of aldosterone secretion.
Ø A series of nitrogen compounds like nitroprusside, nitroglycerine, nitric oxide (NO), sodium nitrite and sodium azide that are known to cause smooth muscle relaxation and vasodilation are also known to increase the activity of cytosolic guanylyl cyclase and increase cGMP levels to exert these effects.
Ø cGMP in turn binds to and activates cGMP – dependent protein kinase which in turn phosphorylates a number of smooth muscle proteins including myosin light chain thereby causing smooth muscle relaxation and vasodilation.
Ø Inhibitors of cGMP – phosphodiesterase enhance the effects brought about by cGMP and prolong these responses for more time. E.g. sildenafil.
VISION,
one of the functions classified under special senses, is carried out by the
photo-sensitive receptors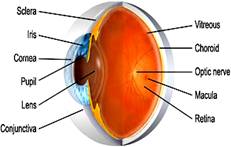 of the retina present in the eyes.
of the retina present in the eyes.
RETINA: The vertebrate retina is a thin sheet of tissue lining the posterior part of the eye. It is highly organized and consists of several layers of neurons. This is the inner nervous layer of the eye where inverted cgm of the objects is formed. It develops from primate brain and has following main layers from within outwards:-
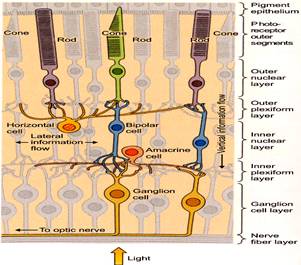
1. LAYER OF OPTIC NERVE FIBRES: Formed by axons of the ganglion cells, which converge to form the optic nerve.
2. LAYER OF GANGLION CELLS: They have pyriform ganglion cells of various sizes.
3. INNER PLEXIFORM LAYER: Has synapses formed by dendrites of ganglion cells and axons of bipolar cells.
4. INNER NUCLEAR LAYER: Mainly formed by bipolar cells which are small and oval-shaped.
5. OUTER PLEXIFORM LAYER: Has synapses formed by dendrites of bipolar cells and axons of rods and cones.
6. OUTER NUCLEAR LAYER: Formed by fibres and nuclei of rods and cones.
7. LAYER OF RODS AND CONES: It is the photo-receptor layer. Rods are cylindrical, have rhodopsin pigment and are associated with dim light-vision (scotopic vision). Cones are pyramidal-shaped and are associated with bright light (photopic vision) and colour vision.
8. PIGMENT CELLS LAYER: Composed of single layer of hexagonal epithelial cells that have a dark brown pigment (fuchsin).
PATH OF LIGHT
Light has to traverse ganglion and bipolar cells to reach the hν-receptor rods and cones. Visual impulses are set up there that are passed on to bipolar cells of inner nuclear layer and then to ganglion cells. The ganglionic axons that form the optic nerve fibre pass the impulse on to cerebral cortex (occipital lobe).
BLIND SPOT: It is the spot at the back of the eye from where the optic nerve fibres leave. This region is covered by a zone of retina free from rods and cones. So, this spot is devoid of the ability for vision.
FOVEA: Lateral to the blind spot, there is a depressed area of the retina called fovea. This site contains only the cones and no rods. The ability for vision is highest in fovea. When the eyes are fixed on an object, its cgm is focused on the fovea and is consequently seen most accurately.
THE PHOTO-RECEPTORS
F Rods are exquisitely sensitive to light and can even detect single photons.
F They allow vision in dim light.
F In contrast, cones are less (30-100-fold) sensitive to light than rods.
F They are used for vision during day light and for colour vision.
F The highest density of cones is found in the center of the retina, the area centralis.
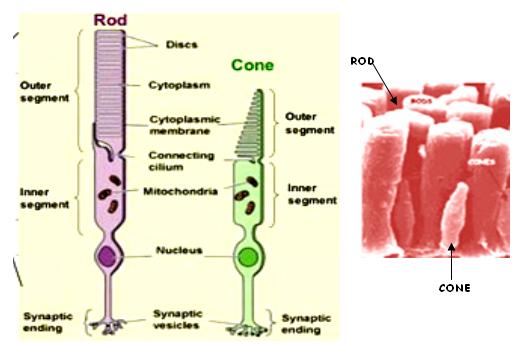
F Both cell types, rods and cones, consist of 4 major parts: -
1. AN OUTER SEGMENT: contains the molecular components for light absorption and generation of electrical signals.
2. AN INNER SEGMENT: harbours the machinery for protein synthesis and energy production.
3. A thin rudimentary CILIUM that connects the above two parts, and
4. A SOMA AND AN AXON with a synaptic region.
THE PHOTO-SENSITIVE COMPOUNDS
In humans and most other mammals, OPSIN (a protein) and RETINENE1, (the aldehyde form of Vit-A1), constitute the photo-sensitive compounds. Retinene is also called as retinal (-CHO).
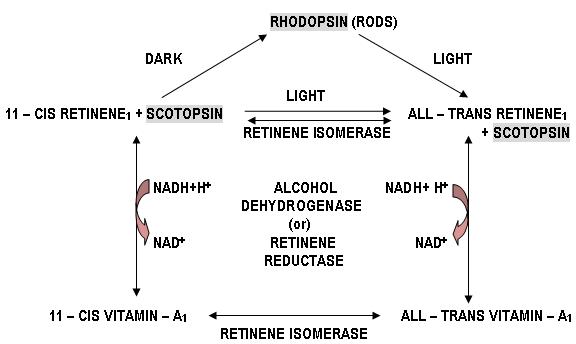
RHODOPSIN, present in the rods, is also known as visual-purple. Its Opsin is called Scotopsin. Rhodopsin has a peak sensitivity to light at λ = 505 nm. Human rhodopsin is a purple-red protein with mol. wt. 41 kD. It is found in the rod-disk membranes and makes up to 90% of the total proteins there. It is one of the G-Protein Coupled Receptors. Retinene1 is attached to 3 of the 7- transmembrane domains parallel to the membrane surface.
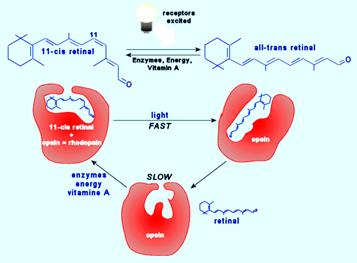
PHOTOCHEMICAL & BIOCHEMICAL CHANGES IN VISUAL RESPONSE – WALD’S VISUAL CYCLE
1. Rhodopsin, which is the chromoprotein that is present in the outer segment of the rods, is bleached on exposure to light.
2. When light falls on retina, the rhodopsin passes rapidly through several stages of very short half – lives.
3. It is finally converted to metarhodopsin – II that splits in to all – trans retinene1 and opsin.
4. This all – trans retinene1 is then reduced to vitamin – A1 by alcohol dehydrogenase (otherwise called retinene reductase) using NADH (note that usually NADPH serves as coenzyme for reductase enzymes but, here is one of the exceptions). It is then stored in the pigment epithelium.
5. Similarly, 11 – cis retinene1 can be converted to 11 – cis vitamin – A1, by alcohol dehydrogenase. These two are reversible reactions.
6. The photopigment should be regenerated after being bleached so as to get ready for the next round of response.
7. Therefore in the dark, retinene isomerase and also vitamin – A1 isomerase, converts all – trans retinene1 and all – trans vitamin – A1, to 11 – cis retinene1 and 11 – cis vitamin – A1, respectively. These two are also reversible reactions. This reaction consumes energy and takes place in light of short wavelength.
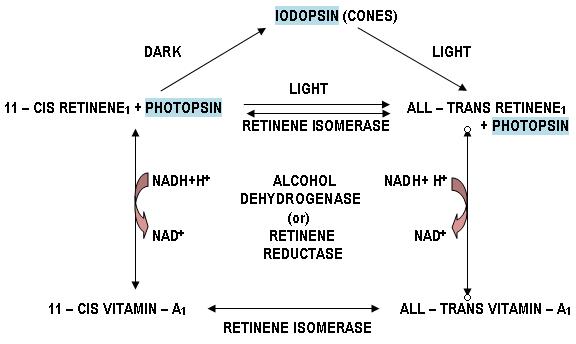
8. Similar changes takes place in cones where instead of rhodopsin we have iodopsin which splits up to photopsin and all – trans retinene1.
9. The trans form of vitamin – A (retinol), which is abundant in the liver and blood, is the most active isomer in all physiological roles of the vitamin, except vision.
10. Bleaching and regeneration of photosensitive pigments are independent of nerves but takes place only as long as the rods remain in contact with the stratum pigmenti.
11. Bleaching itself or products formed during bleaching stimulate the rods to generate visual impulses, i.e., light energy is converted to nerve impulse.

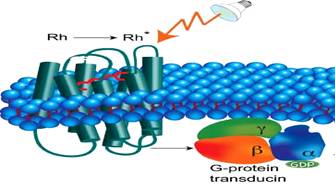
ROLE OF GPCR IN VISION
In photoreceptor cells, Giαt, (belonging to Gi class of G-proteins) causes increase in cGMP PDE activity on stimulating with light. This gives rise to vision.
FUNCTION OF cGMP
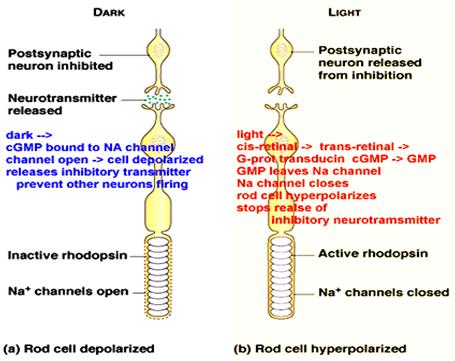
1. In the dark, the intracellular concentration of cGMP is high.
2. The second messenger binds to ion channels in the plasma membrane of the photoreceptor cell.
3. These channels are directly gated open by cGMP and are therefore, referred to as cyclic nucleotide-gated (CNG) ion channels.
4. Few percent of CNG channels are kept open in the dark by binding of cGMP and allow the influx of ions into the outer segment.
5. This influx is primarily carried by sodium (Na+) ions, but a small fraction is also carried by Ca2+ ions.
6. At the same time K+ ions leave the inner segment through potassium (K+) channels.
7. Due to this circulating current along the rod photoreceptor, the so-called dark-current, the membrane potential of the photoeceptor cell is slightly depolarized. This depolarization suffices to release the neurotransmitter glutamate from the synaptic terminal onto postsynaptic cells in the retina, the horizontal and bipolar cells.
8. Upon illumination, the cGMP is hydrolyzed by the PDE.
9. The cGMP concentration decreases and CNG channels close.
10. Since current stops flowing into the cell, the cell hyperpolarizes and glutamate release ceases.
11. The change in neurotransmitter release is registered by bipolar cells, which relay this information onto ganglion cells. The ganglion cells then convey the information to the brain.
RECOVERY OF PHOTO-RECEPTORS
Recovery of the photoreceptor from the light response and adjustment of light sensitivity is achieved by several feedback mechanisms:-
1. Ca2+ ions play a major role in recovery and light adaptation.
2. Upon illumination, Ca2+ entry through the CNG channel ceases, whereas a Na+/Ca2+-K+ exchanger continues to export Ca2+ ions from the cell.
3. The ensuing decrease of the intracellular Ca2+ concentration triggers recovery and adaptation.
4. Ca2+ controls the synthesis of cGMP by guanylate cyclase (GC).
5. The enzymatic activity is controlled by specific Ca2+-binding proteins (GCAP's). Furthermore, inactivation of enzymatically active rhodopsin is achieved by phosphorylation through Rhodopsin Kinase.
6. This process involves another Ca2+-binding protein, Recoverin.
7. Finally, the ligand sensitivity of the CNG channel is controlled by a third Ca2+-binding protein, Calmodulin.
NEUROTRANSMITTERS IN THE RETINA
Acetyl choline (Ach), dopamine, serotonin, γ – Amino butyric acid (GABA), glycine, substance P, glutamate and aspartate are some of the chemical transmitters present in the retina. Also, somatostatin, glucagon, neurotensin, CCK, VIP, TRH, LRH, encephalins, β – endorphins are present.
LIGHT & DARK ADAPTATIONS
On either moving from a darker area to brighter or vice versa, our eyes take a little time to get adapted to the situation. These two are called dark adaptation (scotopic eye) and light adaptation (photopic eye).
DARK ADAPTATION: On moving from a brightly illuminated area to a dark area, the vision is poor at first, but improves after some time. This is dark adaptation. It is completed within 1 hour. The time taken for dark adaptation is directly proportional to the intensity of light and duration of exposure and inversely proportional to vitamin – A content.

Both cones and rods take part in dark adaptation. Cones adapt earlier than rods. At first, sensitivity of vision rises quickly to reach a fairly steady value within 4 – 5 min and it is maintained for another 5 – 6 min. This is called cone adaptation. After that the sensitivity is increased, but slowly, with much greater degree. This is called rod adaptation.
LIGHT ADAPTATION: On moving from a dark area to a brightly illuminated area, we feel intense and uncomfortable light. But, after 5 min, this stage passes over and the visual threshold of the eye rises. This is called light adaptation. Light adaptation is simply the disappearance of dark adaptation.
DUPLICITY THEORY OF RETINAL FUNCTION (OR) DUPLEX THEORY OF VISION - SPECTRAL SENSITIVITY: SCOTOPIC AND PHOTOPIC VISIONS
According to this universally accepted theory of vision, the cones are responsible for (a) bright light vision, (b) visual acuity and (c) color vision. But, the rods are responsible for dim light vision, i.e., objects appear only in shades of gray and no other details or color of the object are seen.
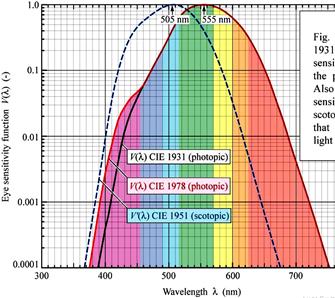 LUMINOSITY
CURVE: A graph plotted by taking
luminosity along y-axis and wavelength along x-axis. This graph shows the
relative luminosity of various wavelengths of light. In
scotopic vision,
maximum luminosity is observed at
507 nm (blue – green) and in
photopic vision,
maximum luminosity is observed at
550 nm (yellow – green).
LUMINOSITY
CURVE: A graph plotted by taking
luminosity along y-axis and wavelength along x-axis. This graph shows the
relative luminosity of various wavelengths of light. In
scotopic vision,
maximum luminosity is observed at
507 nm (blue – green) and in
photopic vision,
maximum luminosity is observed at
550 nm (yellow – green).
This shift of spectral sensitivity towards longer wavelength is known as Purkinje shift.
This phenomenon can be observed by looking at red and blue flowers in a garden in the evening. When light is still present, both of them appear bright. But, as light fades, the red flower appears dark, while the blue flower appears to be coloured.
COLOUR VISION
The sensitivity of the eye to a flash of light varies with the wavelength of the light. The color of light depends upon its wavelength and the visible spectrum extends from 397 – 750 nm (VIBGYOR). Beyond this range, the photoreceptors are not stimulated. The normal human eye can detect almost all the gradations of colour as many as 160. This means that the retina can detect differences of about 3 nm.
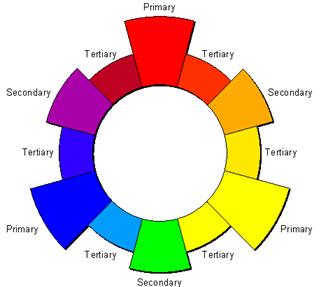
1. PRIMARY COLOURS: Red, Green and Blue are primary (1o) colours. These are called primary because, by mixing them in suitable proportions, any desired colour can be obtained.
2. SECONDARY COLOURS: Mixing any two 1o colours will give rise to secondary (2o) colours. Red + yellow = orange; yellow + blue = green and blue + red = purple.
3. TERTIARY COLOURS: Mixing the 1o colours with their adjacent 2o colours results in six tertiary (3o) hues. Red-orange, yellow-orange, yellow-green, blue-green, blue-purple and red-purple.
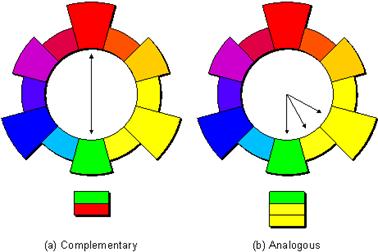
4. COMPLEMENTARY COLOURS: These are any two colors that are directly opposite each other on the color wheel and provide maximum contrast, such as red and green, yellow and blue, etc.
5. ANALOGOUS COLOURS: These are any three colors that are side-by-side on the color wheel, such as yellow, yellow-green, and green.
WAVELENGTHS AND PAINTS MIX IN A DIFFERENT MANNER
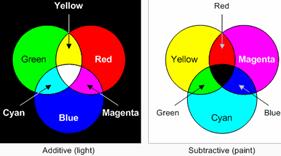
When talking about mixing of two or more colours that in turn produce a different colour sensation, it should be kept in mind that, only wavelengths of the colours mix with each other. Colour paints or colour pigments do not mix in the same manner. E.g.: Yellow + blue light = white light, but, yellow + blue paint = green paint. This is because, blue paint absorbs all light except blue and a little green and yellow paint absorbs all light except yellow and a little green, so we see green paint. This is true for all coloured substances that we see.
GRASS IS GREEN – YES OR NO?
Mixing of wavelength is additive, while mixing of paints or pigments is subtractive.
The red paint absorbs the green and blue light, but it reflects the red light, which is what we end up seeing. Similarly, the green paint absorbs the red and blue light and reflects the green, while the blue paint absorbs the red and green and reflects the blue. The white paint reflects all of the colors and the black paint absorbs them all, which means that black is really an absence of any color.
When we look at the grass, what we actually see are the colors it didn't absorb. We could say that grass is green because that's the color that it reflects for us to see, or we could say that grass is both blue and red because those are the colors it absorbs.
This explains why mixing paints are different from mixing light. If we start off with two tins of paint – say cyan and yellow – and shine white light at them, then each of the paints absorbs some of the colors from the white light and reflects others. If we now mix the two paints together, they each continue to absorb the same colors that they did before, so we end up seeing whichever colors neither of them absorbed, which is green in this case. This is why we say mixing paints is subtractive, because the more paints we mix together, the greater the number of colors the combination subtracts from the white light.
Red, Green and Blue are called additive primary colours, whereas, Cyan, Magenta and Yellow are called subtractive primary colours. (See diagram above)
THE 3 ATTRIBUTES OF COLOUR
a. HUE: This depends entirely on the wavelength of light. VBGYOR are the hues. Indigo is removed from the spectrum list.
b. BRIGHTNESS: Depends on the intensity of rays and sensation.
c. SATURATION (OR) PURITY: It varies with the amount of white mixed with it. E.g., red is more pure than pink (pink = red + white).
The sensation produced by a colour also depends on the colour of the surroundings. E.g.: red is seen as red when it has green or blue background. But, it appears pale pink on a red background. This is called contrast phenomenon.
THEORIES OF COLOUR VISION
Many theories were put forth to explain how we perceive colour. But, none of them is wholly perfect.
1. YOUNG – HELMHOLTZ THEORY: This is also known as the trichromatic theory of colour vision. It was proposed by Thomas Young (1801) and in the middle of the same century it was further detailed by Helmholtz. This is the widely accepted theory and holds good when it comes to explain colour vision defects.
According to this theory, there are 3 types of cones for the 3 basic colours (RGB) each containing a different photochemical substance. Each photoreceptor and photopigment is specifically acted upon by light of the corresponding hue (λ), while other colours have lesser or no effect. E.g.: red light stimulates red – sensitive cones maximally and cause decomposition of the red pigment. It will stimulate green and blue cones only feebly.
Sensations of various colours and white are due to stimulation of different receptors at different intensities. The impulses set up are conducted along separate nerve fibres to different neurons in the visual cortex.
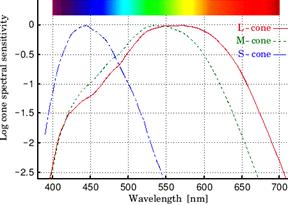
Absorption spectral studies in man have shown the existence of 3 types of cones, each having a different photochemical substance, namely:
a. S – CONE (or) SHORT WAVELENGTH CONE: The pigment whose λmax is 440 nm (430 – 460 nm) that senses blue colour. It is called cyanolab.
b. M – CONE (or) MEDIUM WAVELENGTH CONE: The pigment whose λmax is 535 nm (520 – 545 nm) that senses green colour. It is called chlorolab.
c. L – CONE (or) LONG WAVELENGTH CONE: The pigment whose λmax is 570 nm (560 – 585 nm) that senses red colour. It is called erythrolab.
The centre of fovea is blue – blind because it does not have blue cones in that region.
2. GRANIT’S POLYCHROMATIC THEORY OF COLOUR VISION (or) DOMINATOR – MODULATOR THEORY: This theory was proposed by Granit (1945). He states that:-
a. Ganglions called dominators are stimulated by the whole of visible spectrum.
b. There are 2 types of dominators: a) dominator for rod – these respond only in dark – adapted eye (scotopic) and give maximum response at 500 nm; b) dominator for cone – these respond in light – adapted eye (photopic) and give maximum response at 550 nm.
c. Dominator rod is related to brightness (luminosity) and dominator cone is related to intensity of light and not colour.
d. In light adapted eye, certain ganglions called modulators are stimulated only at a narrow wavelength band.
e. Modulators are of 3 groups: a) affected by blue light (450 – 470 nm); b) affected by green light (520 – 540 nm) and c) affected by red – yellow light (580 – 600 nm). Thus, they are concerned with colour sensation.
f. If green light falls on the retina, it stimulates the green modulator maximally, while blue and red modulators are affected little. Similar for other colours.
3. HERING’S OPPONENT COLOUR THEORY OF VISION: This theory was proposed by Ewald Hering (1964). It is a modification of Young’s theory. Hering also assumes the existence of 3 types of photochemical substances in the retina but, he states that, they tend to give 6 different qualities of colour sensations.
|
PHOTOCHEMICAL SUBSTANCE |
RETINAL PROCESS |
SENSATION |
|
White – Black |
Breakdown |
White |
|
|
Resynthesis |
Black |
|
Red – Green |
Breakdown |
Red |
|
|
Resynthesis |
Green |
|
Yellow – Blue |
Breakdown |
Yellow |
|
|
Resynthesis |
Blue |
According to this theory, the photochemical substances (complementary colours) give one sensation on breakdown (decomposition) and a different sensation on resynthesis. So, the complementary colours become antagonistic to their respective primary colours.
Hering does not agree the presence of separate nerve fibres for different colour sensations. He assumes that the same nerve fibre can carry impulses from two different sensations to the brain.
COLOUR BLINDNESS
Colour blindness means defect or lack of appreciation of one or more colours. It is of many types and can be classified according to Young’s theory like:
1. ANOMALOUS TRICHROMATISM: This is the commonest form. These people can see all the 3 colours, but the intensity to identify a particular colour should be more when compared to a normal person. It has 3 subtypes:-
a. PROTANOMALY (PARTIAL PROTANOPIA): Red vision is subnormal. It is generally congenital.
b. DEUTERANOMALY (PARTIAL DEUTERANOPIA): Green vision is subnormal. It is generally congenital. It is very common of the 3 subtypes.
c. TRITANOMALY (PARTIAL TRITANOPIA): Blue vision is subnormal. It is the rarest of the 3 subtypes.
2. DICHROMATISM: This is much less common than the first type. These people can see any 2 colours only. Not all the 3 colours.
a. PROTANOPIA (red blindness). It is congenital.
b. DEUTERANOPIA (green blindness). It is congenital.
c. TRITANOPIA (blue blindness). It is usually acquired and it is very rare.
3. MONOCHROMATISM: The colour sensation of the cones is lost. The rods are normal. It is called total colour blindness. In day time, the surrounding appears to be dark, grey or pale bluish – white (scotopic).
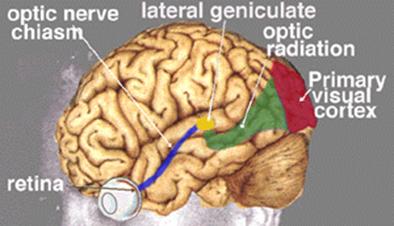
VISUAL IMPULSE TRANSMISSION PATHWAY TO BRAIN
cGMP – PHOSPHODIESTERASE (cGMP-PDE):
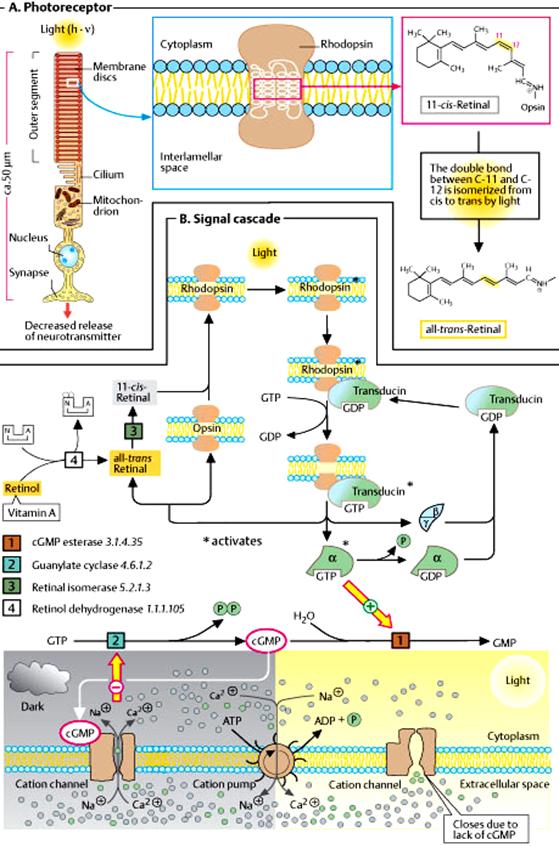
Actions that are caused by hormones that increase cAMP or cGMP levels, can be stopped in a number of ways, including hydrolysis of cAMP/cGMP by phosphodiesterases (PDE). During the presence of light, i.e., bleaching of photoreceptors, the following events take place:-
1. Rhodopsin converts to metarhodopsin – II in a series of steps.
2. This change in the GPCR namely rhodopsin, induces the Giαt (Transducin).
3. The α subunit of transducin splits off from the β,γ – subunits.
4. Then GDP is removed from α – subunit and GTP is added to it to make it active.
5. This α – GTP complex induces the enzyme cGMP – PDE which catalyzes the hydrolysis of cGMP to GMP.
6. Due to the absence or very low level of cGMP, the cyclic nucleotide-gated (CNG) ion channels present in the rod cell membrane closes.
7. So, Na+ channels close and no Na+ influx is stopped which will make the cell hyperpolarized (outside more + ve inside more – ve).
8. Hyperpolarization causes the inhibition of release of the inhibitory neurotransmitter namely glutamate.
9. Since glutamate is not produced nerve impulse transmission takes place and the sensation of light is felt. (refer diagram in unit-IV page 8)
Inhibitors of cGMP – PDE will cause an increased level of cGMP and prolong the actions mediated by it. These play an important role in relaxation of smooth muscle and vasodilation.